microRNA(miRNA)常以miRNA沉默复合体(miRISC)的形式,结合于mRNA的3’UTR区域,参与转录后调控。最近,有研究发现,miRNA也可以通过外泌体分泌到胞外,直接作为配体,与Toll样受体(TLR)互作,产生不同信号通路,发挥截然不同的作用。
来自德国柏林夏里特大学的研究者在近期研究中发现,一种let-7家族的miRNA:let-7b会进入皮层神经元中,利用一个多GU序列,结合并激活细胞核内体中的TLR7,促使TLR7富集接头蛋白MyD88,产生细胞凋亡反应,引起神经退行性病变。这个过程可以概括为let-7b/TLR7/MyD88通路。在此通路中,let-7b可能来自其他濒死神经元分泌的外泌体,也有可能来自细胞自身分泌外泌体。let-7b的这种功能,可能会引起神经性变相关疾病发生,如阿尔茨海默病等。
然而,在大洋彼岸的美国,杜克大学研究者却得到了与德国同行不同的研究结果。他们发现,在与疼痛感知相关的背根神经节(DRG,痛觉传入的第一级神经元,在痛觉的外周机制中起重要作用)神经元受到伤害性刺激时,细胞会上调并释放let-7b,与细胞膜上的TLR7互作,激活一个阳离子通道瞬时感应器A1(TRPA1),通过细胞表面电位变化,产生疼痛过程,这个过程不需要髓鞘,而且是缓慢发生的。
两所大学的研究者在对神经元中miRNA进行研究时,发现了不同的功能。在皮层神经元,let-7b这个miRNA会诱发细胞凋亡,引起神经退行性疾病;但在DRG神经元中,let-7b却又会刺激TLR7受体,提高神经元细胞敏感度。这种功能差异是如何产生的?
第一类解释认为,是不同神经细胞中TLR7定位不同造成了let-7b的功能差异。在皮质神经元中,转运蛋白UNC93B1会特异性地将TLR7从内质网经高尔基体运送到核内体中,但在DRG神经元中,TLR7并不会被运送到核内体,而是通过其他途径,定位到质膜上。不同位置的两种TLR7与let-7b互作,产生信号。除了转运蛋白,转运过程中其他因素也可以影响TLR7定位,如pH、辅助因子等。而第二类解释则认为,是不同的配基,让不同神经元中的let-7-TLR7互作产生了不同信号。在皮质神经元中,let-7b利用一个富含GU的序列与TLR7结合,而这个序列如果稍作改变,还可以结合其他RNA,产生不同信号,在其他let-7家族成员刺激TLR-7时就能发现这种现象。所以,这种解释认为,是不同的配体,让let-7-TLR7互作产生了不同的信号。
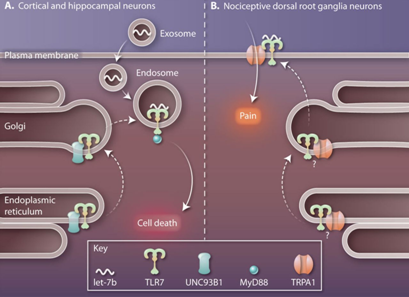
图1. 不同神经元中let-7b-TLR7互作通路(A)在皮层神经元中,UNC93B1蛋白将TLR7由内质网经高尔基体定位到核内体中,神经元吸收含有let-7b的外泌体后,let-7b与核内体中的TLR7互作,进一步富集MyD88蛋白,促使神经元凋亡(B)DRG神经元中,TLR7经其他通路定位到细胞膜上,神经元在受到伤害性刺激时,上调并分泌let-7b,let-7b与细胞膜上TLR7互作,影响TRPA1,产生细胞膜电位变化,引起疼痛反应。
以上这些研究内容,可以说颠覆了miRNA的传统作用模式,miRNA不再以miRISC的形式参与mRNA转录后调控,而是与受体直接互作,产生信号。一方面,以let-7b为代表的let-7家族miRNA会通过外泌体进入皮层神经元细胞,激活核内体中的TLR7,促使细胞凋亡发生,诱发阿尔茨海默及其他神经变性相关疾病;另一方面,在疼痛DRG神经元中,let-7b又可以与细胞膜上的TLR7互作,进一步激活TRPA1,改变细胞膜电位,产生疼痛反馈,提高神经元细胞灵敏度。这些研究,扩宽了miRNA和TLR7在神经反应过程中的调控作用,并证明TLR7在胞内的不同定位可能促成了神经元之间功能差异。确定TLR7在不同神经元中的定位、转运过程以及各个信号通路中的信号分子,可以更好地理解miRNA是如何调控中枢神经系统(CNS)的健康和疾病状态。同时,通过了解miRNA的来源,可以针对miRNA进行靶向作用,影响miRNA功能,也可以直接利用外泌体miRNA与不同受体结合,产生响应信号,这可能成为治疗神经疾病的新思路。
Location Is Everything: let-7b microRNA and TLR7 SignalingResults in a Painful TRP
Extracellular let-7b, a microRNA found in the central nervous system, affects neurons through its interaction with Toll-like receptor 7 (TLR7), but with divergent outcomes in different neurons. Lehmann et al. found that let-7b stimulation of cortical and hippocampal neurons led to neuronal apoptosis, whereas Park et al. report that let-7b activation of TLR7 stimulated the cation channel transient receptor potential A1 (TRPA1) on dorsal root ganglia sensory neurons and induced pain responses. The primary difference that may infl uence these distinct responses to let-7b is the localization of TLR7 to the endosome in the cortical and hippocampal neurons or the plasma membrane in the sensory neurons. These studies suggest that different types of neurons traffi c TLR7 to distinct membrane locations, affecting the functional response of neurons to let-7b stimulation.